Aspects of Functional Morphology
When cephalopods were first found with aptychi in place, the approximate match between their outline and the aperture of the conch suggested they served as opercula. This correspondence is by no means exact, however (Lehmann, 1972, p.42). Other functions postulated for these structures were as covers for the nidamentary glands or as cartilaginous plates for funnel muscle attachment. Ruedemann (1916) suggested the latter, "would naturally also have existed in the Ordovician and Silurian cephalopods..." in attempting to explain the Discinocarida.
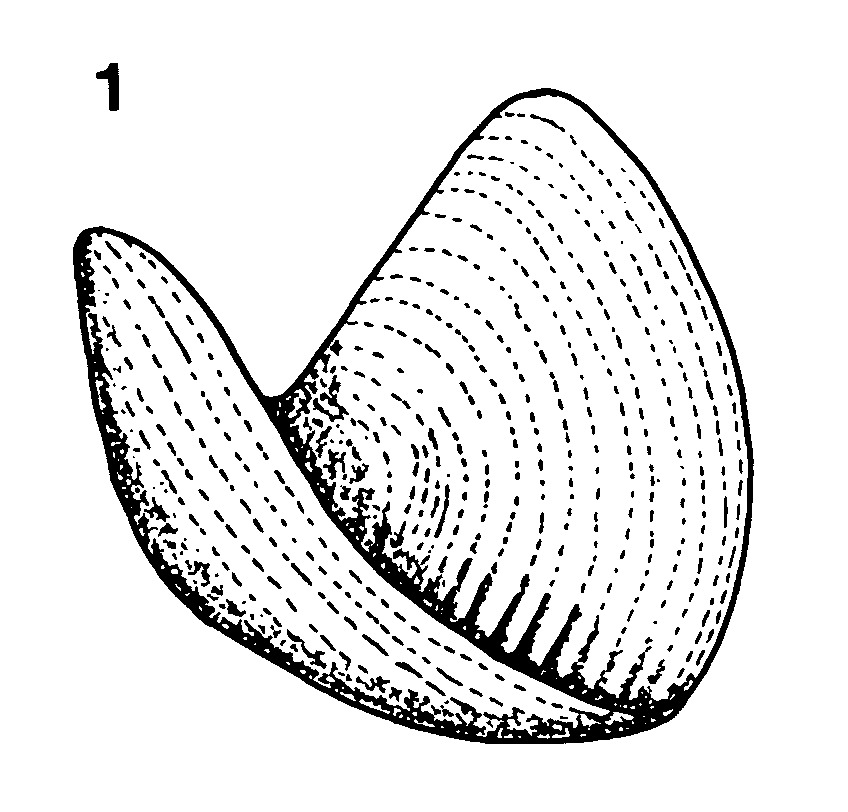
This prescient speculation was proven sound by the discovery of specimens of Aptychopsis Barrande, 1872 in situ in the apertures of orthoconic nautiloids from the Silurian of central Bohemia (Turek, 1978) and southern Sweden (Holland, et al., 1978). Aptychopsid plates form a neat, flat circular structure almost precisely fitting the aperture of the nautiloid (Stridsberg, 1984). The three plates involved would be difficult to fold into a concave structure (Fig. 14.1). It would seem that these are indeed opercula.
1.  2.
2. 
3. 
Figure 14 Aptychopsis Barrande, 1872. 1. LO 5270 and 2. LO 5268, from a Silurian shale quarry in southern Sweden (Stidsberg, 1984). Specimens from the Department of Historical Geology and Palaeontology, University of Lund, Sweden. 3. Reconstruction of opercular position of aptychopsid plates in an orthoconic nautiloid, after Turek (1978).
The structures known as aptychi in Mesozoic ammonites seem to have served a different function. Again, discoveries of aptychi in situ in cephalopod body chambers provided evidence of their function. These were found to be curved structures, sometimes found associated with an element resembling the upper jaw of Nautilus (Lehmann, 1971; 1972; 1978; 1981; Tanabe, 1983; Tanabe and Fukuda, 1987). It has become accepted by many that these aptychi are the lower jaws of ammonites (Lehmann, 1981; Morton, 1981; Mapes, 1987). One difficulty with this interpretation is the sheer size of the structure, relative to both the upper jaw components and the diameter of the body chamber. In many cases, the aptychus approximates the cross-sectional area of the body chamber. Lehmann's work, cited above, involves serial sectioning of cephalopods, revealing the three-dimensional relationship of the body chamber and its contents, and seems to be the most accurate means of determining the nature of these structures, at least for the Mesozoic ammonites studied.
There are no rigid calcareous plates found with the Devonian fossils, so their preservation as flattened bodies does not necessarily reflect their original morphology. Two specimens of Sidetes newberryi, CMNH 8311 and CMNH 8318, exhibit short cracks radiating inward from the posterolateral margin, which are visible in Figure 8.2. These cracks are just what would be expected in a convex structure which has been flattened. If this is the origin of the cracks, then closing the gaps should approximate the original form of the structure. As these specimens were folded in half along the medial line, photographs of them were prepared in both normal and reversed orientations. These were then photocopied and enlarged. The resulting reproductions were then cut out and the mirror images were attached along the midline. This caused the paper models to become slightly convex. The cracks along the margin were then cut away and the edges rejoined. This caused the models to assume a broad, scooped shape, similar to that seen in Lehmann's (1970) reconstruction of the jaws of Psiloceras from the Jurassic of Germany (Fig. 15). Many of the specimens preserved as flattened bodies are folded along the median. In some cases they can be separated from the matrix and studied from both sides, or often the remains of the upper half are preserved along the margins of the extant lower portion (see the arrow in Fig. 4.2, S. emersoni).
{kind=link}
1.  2.
2. 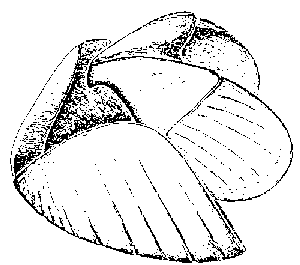
Figure 15 Hypothesized original shapes of aptychi. 1. Reconstruction of a Cleveland Shale anaptychus. 2. Reconstruction of the mandibles of Psiloceras, Hyatt 1867 (from Lehmann, 1971) Click on the thumbnails for enlargements.
These reconstructions of ammonite jaws (Lehmann, 1970; 1975), suggest to some a scooping, shoveling application rather than a true biting action seen in modern Nautilus (Lehmann, 1972), and in other Mesozoic forms (Nixon, 1988). This conclusion is based on the large size of the lower jaws (particularly anaptychi), both relative to the size of the upper jaw and to the total size of the conch. The lack of calcified rostra, or conchorhynchs, to serve as biting surfaces further serves as a basis of interpretation. Calcified beaks have been reported from the Permian, (Closs, et al., 1967) and perhaps from the Mississippian (Landman and Davis, 1988), but no older specimens have been found. Where associated upper jaws are of comparable scale, however, it would seem that, even lacking calcified beaks, the jaws could be capable of strong biting action similar to modern coleoids, which lack such calcified surfaces (Tanabe, et al., 1980). The shape of the lower jaw is similar for both applications. Still, the curved shape revealed in the Ohio Shale specimens by these methods, if genuine, fits known jaw structures far better than hypothesized opercula or Aptychopsis. We must conclude that these forms of Sidetes from the Late Devonian represent cephalopod jaws which have been preserved separately from the animal's conch.